导读:拉马克认为生物在新环境的直接影响下,习性改变,某些经常使用的器官发达增大,不经常使用的器官则逐渐退化(用进废退),并认为这样获得的后天性状可传给后代,生物体由此可逐渐演变。此外,他还认为适应是生物进化的主要过程。

长颈鹿脖子的用进废退进化
 达尔文认为遗传变异和自然选择决定物种由简单到复杂,由低等到高等的进化,并提出著名的“物竞天择,适者生存”的论断。他还认为遗传突变是生物进化的动力,有利突变可在自然选择中被保存。这种进化论的着眼点是群体,遗传物质的多样性通过个体的遗传突变而扩增,进而能够更好地适应环境。
达尔文认为遗传变异和自然选择决定物种由简单到复杂,由低等到高等的进化,并提出著名的“物竞天择,适者生存”的论断。他还认为遗传突变是生物进化的动力,有利突变可在自然选择中被保存。这种进化论的着眼点是群体,遗传物质的多样性通过个体的遗传突变而扩增,进而能够更好地适应环境。

随着遗传物质的测序和中心法则的认识,越来越多的人相信达尔文进化论是科学的,个体意识能支配进化的拉马克主义只是谬论。“物种漂变学说”和“中性进化”都是达尔文主义的现代版,它们解释了遗传物质—基因在物种进化中的演变。
随着科学的向前发展,有些生物现象不能够被达尔文学说解释。如空气中的污染物在没有改变基因的情况下如何影响DNA的表达,从而导致像肺气肿或肺癌之类的疾病;某种疾病只发生在同卵双胞胎中的一位。这些现象排除了遗传物质变异的可能性,而生物特性却出现改变。表观遗传学是遗传物质之外的遗传因素,也是外界环境与遗传物质之间的桥梁。在遗传物质没有改变的情况下,生物体的性状受到特定环境诱导而发生可遗传的变化。从性状遗传上看,表观遗传学是拉马克学说的延续,生物体性状在物种繁衍过程中不断被继承。一些组装信息(如DNA甲基化和染色质构象变化等)可调控基因的表达水平,进而影响节遗传基因的功能和特性。
然而表观遗传学克服了生物体进化的唯意识论,因而不是简单的拉马克学说。不仅没有否认达尔文的进化论,它还作为这一理论的补充。生物体性状的强化或退化不是生物体的意识决定的,而是受遗传物质和表达模式共同影响的。关于生物的进化,达尔文从遗传物质解释,而表观遗传学则从基因的表达模式上诠释。换句话说,达尔文回答了人类是由古猿进化的,而表观遗传学则回答为什么一方水土养育一方人。

 表观遗传“记忆”
表观遗传“记忆”
约翰英纳斯中心的研究人员发现了生物体对不同环境条件(如营养的质量或温度)形成记忆的证据,这一发现解释了记忆机制和记忆如何遗传给下一代。
基因的表达活性在一些地区长期受环境的影响;个体生存的环境能影响后代的生物学或生理学特征,但是在遗传过程中基因组却没有改变。一些研究表明,在家庭中,祖父母遭受严重的食物短缺,子孙后代患心血管疾病和糖尿病的风险就会更大,表观遗传学中通过基因是否表达获取的记忆能解释这一点。但是截止到目前,还没有一个清晰的机制能解释个体如何形成对可变因子(如营养)的“记忆”。
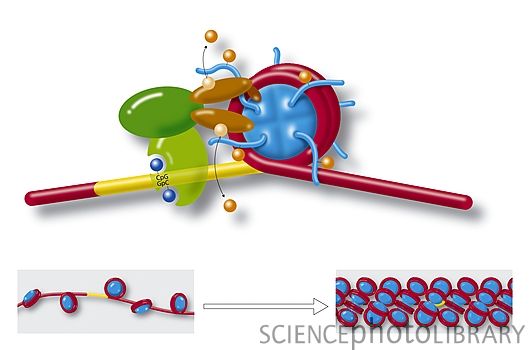
表观遗传学中甲基化对基因表达的作用
研究表明组蛋白甲基转移酶和去甲基酶对于组蛋白甲基化状态影响很大,比如H3K9特异性去甲基化酶Jhdm2a在激素受体介导的基因激活中扮演了重要角色。
Jhdm2a在调控代谢基因表达方面发挥了重要作用。缺乏Jhdm2a会导致细胞中β-肾上腺素刺激的糖释放和棕色脂肪组织中氧消耗的紊乱,以及降低骨骼肌肉中脂肪氧化和糖释放。
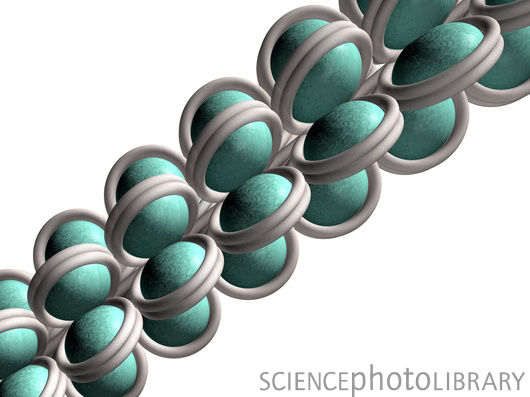
表观遗传学的组蛋白修饰
当精子遇上卵细胞时,受精卵首先要清除来自亲代的特殊程序信息(精子或卵细胞自身的遗传程序信息),然后再开始生长成为一个新的生命。也正是这个原因,子代与亲代的表观遗传信息发生了深刻的变化。
程序重排与一个特殊的组蛋白修饰作用有关,如果这个组蛋白修饰发生突变将导致清除作用(清除亲代的表观遗传学信息)失效。正常的C.elegans繁殖下一代的时候组蛋白的修饰信息不会传递给下一代。而如果发生突变,组蛋白的修饰信息会一代代传下去,研究小组做了个实验,突变spr-5(这一突变会导致不孕),同时再突变LSD/KDM1,结果发现子代变得越来越继承亲代的不孕性状。而如果再从外界引进LSD/KDM1又使得子代清除亲代的表观遗传修饰程序的功能得以恢复。
推荐阅读:表观遗传“记忆”








